
The Kelly Group Research
β-Sheet folding:
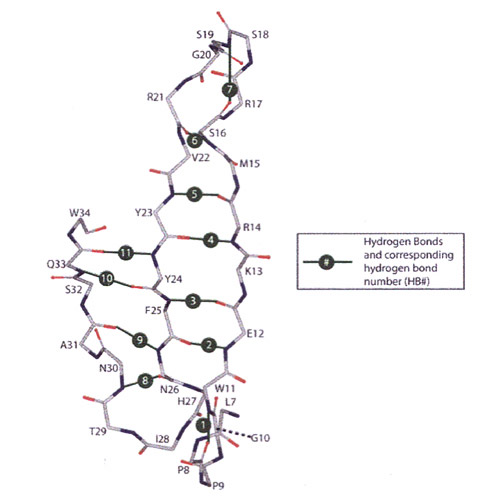
Figure 1: Representation of the backbone of the β-sheet region. Hydrogen bonds and respective hydrogen bond numbers are indicated in green (Deechongkit et al. Toward assessing the position-dependent contributions of backbone hydrogen bonding to β-sheet folding thermodynamics employing amide-to-ester perturbations. J. Am. Chem. Soc. 2004;126:16762-16771).
We use the WW domain of human Pin 1 (Pin WW), a small β-sheet protein comprised of three strands and two loops (Figure 1), as a model system for understanding β-sheet folding energetics. Our kinetic studies, using the laser temperature jump kinetic method in conjunction with Φ-value analysis, revealed that native-like secondary structure exists in the first of Pin WW's two loops in the folding transition state. In addition, we found that the β-strands and the second loop are somewhat structured in the folding transition state, but less so than the first loop. The end of the third strand (the C-terminus of the peptide), however, is almost entirely unstructured in the folding transition state.
The contribution of hydrogen bonding to protein folding kinetics and the stabilization of the folded state continues to be a controversial issue in the protein folding field. Scientists have argued that there are the same number of hydrogen bonds in the denatured state (backbone and side-chain water H-bonds) and the native state, thus in order to be stabilizing a subset of backbone-backbone, backbone-side chain and side chain-side chain H-bonds have to be stronger in the folded ensemble for hydrogen bonding to make a stabilizing contribution. We have used amide-to-ester mutations to evaluate the influence of backbone-backbone hydrogen bonding on the kinetics of protein folding and on the stabilization of the folded state in Pin WW. Amide-to-ester mutations allow us to individually replace backbone amides with esters, thus deleting a hydrogen bond donor (the amide NH becomes an O) and weakening a hydrogen bond acceptor (the amide carbonyl becomes an ester carbonyl). The amide-to-ester perturbations revealed that the Pin WW native state was most destabilized when an amide NH surrounded by hydrophobic residues (preventing aqueous solvent penetration) participating in a hydrogen bond was replaced by an ester oxygen. In other words, this study reveals that only a subset of hydrogen bonds — those buried in the hydrophobic core(s) and excluded from solvent — are important for stability. To assess the strength of individual hydrogen bonds, we perturbed the H-bond between the amide of Phe23 and the carbonyl of Arg14 using an amide-to-ester mutation, which eliminated the hydrogen bond donor and weakened the hydrogen acceptor, or an amide-to-E-olefin mutation, which eliminated both the hydrogen bond donor and acceptor. Comparison of the two mutants enabled us to assign a value of 1.3 kcal/mol to the hydrogen bond.
In a separate study, we further scrutinized the hypothesis that hydrogen bond strengths depend on the polarity of their microenvironment, with stronger hydrogen bonds forming in non-polar surroundings that exclude aqueous solvent. We used the Pin WW described above, and protein A, a largely α-helical protein, as our model proteins. By combining protein backbone and side chain mutagenesis, we demonstrated that decreasing the polarity of the microenvironment of a hydrogen bond can increase its strength by over 1 kcal/mol in both helix- and sheet-rich contexts. This result indicates that accounting for the context dependence of hydrogen bond strengths could be critical for quantitatively understanding any process in which hydrogen bonding plays a central role, including protein folding and ligand binding.
Just as some hydrogen bonds disproportionately contribute to stability of the folded state, so too are some side chain interactions more important than others. A thorough mutational study of the stability of Pin WW was carried out wherein the stabilities of 66 sequence variants of Pin WW were determined by equilibrium thermal denaturation experiments. All 34 residues composing the Pin WW could be replaced one at a time with at least one different natural or non-natural amino acid residue without leading to an unfolded protein. Alanine substitutions at only four positions within Pin WW led to a partially or completely unfolded protein in the absence of a physiological ligand. The side chains of these four residues form a conserved, partially solvent-inaccessible, continuous hydrophobic mini-core comprising the N- and C-termini. Ala mutations at five other residues, three of which constitute the ligand binding patch on the concave side of the beta-sheet, significantly destabilized Pin WW without leading to an unfolded protein. The remaining mutations affected protein stability only slightly, suggesting that only a small subset of side chain interactions within Pin WW are mandatory for acquiring and maintaining a stable, cooperatively folded β-sheet structure.
We have also we evaluated the ability of β-turn mimics to act as nucleators to accelerate β-sheet folding. β-Turns are common conformations that enable proteins to adopt compact globular structures, and their formation is often rate limiting for protein folding. β-Turn mimics, molecules that replace the i + 1 and i + 2 amino acid residues of a β-turn, are envisioned to act as folding nucleators by preorganizing the pendant polypeptide chains, thereby lowering the activation barrier for β-sheet formation. Using six β-turn mimics that have preferred dihedral angle preferences simulating varied β-turn types in place of two residues in an engineered β-turn 1 or β-bulge turn 1 of the Pin WW, we show that inclusion of the β-turn mimics alters β-sheet folding rates, enabling us to classify β-turn mimics into three categories: those that slow but permit Pin WW folding, native-like nucleators, and fast nucleators. This study provides further evidence that the formation of β-turn 1 is rate limiting for WW domain folding and now that these non-natural β-turn mimetics are characterized, they can be used to perturb protein structure in a wide variety of proteins.
- Jager M, Nguyen H, Crane JC, Kelly JW, Gruebele M. The Folding Mechanism of a β-Sheet: The WW Domain. J. Mol. Biol. 2001; 311:373-393.
- Nguyen H, Jager M, Moretto A, Gruebele M, Kelly JW. Tuning the free–energy landscape of a WW domain by temperature, mutation and truncation. Proc. Natl. Acad. Sci. 2003;100:3948-3953.
- Deechongkit S, You S-L, Kelly JW. Synthesis of All Nineteen Appropriately Protected Chiral α-Hydroxy Acid Equivalents of the α-Amino Acids for Boc Solid Phase Depsi-Peptide Synthesis. Org. Lett. 2004; 6:497-500.
- Deechongkit S, Nguyen H, Dawson PE, Gruebele M, Kelly JW. Context Dependent Contributions of Backbone H-Bonding to β-Sheet Folding Energetics. Nature 2004; 430:101-105.
- Deechongkit S, Dawson PE, Kelly JW. Towards Assessing the Position-Dependent Contributions of Backbone Hydrogen Bonding to β-Sheet Folding Thermodynamics Employing Amide-to-Ester Perturbations. J. Am. Chem. Soc. 2004;126:16762-16771.
- Powers ET, Deechongkit S, Kelly JW. Backbone-Backbone H-Bonds Make Context Dependent Contributions to Protein Folding Kinetics and Thermodynamics: Lessons from Amide to Ester Mutations. Adv. Pro. Chem. 2006; 72:39-78.
- Jager M, Zhang Y, Bieschke J, Nguyen H, Dendle M, Bowman ME, Noel JP, Gruebele M, Kelly JW. The Structure–Function–Folding Relationship in a WW Domain. Proc. Natl. Acad. Sci.2006; 103:10648-10653.
- Fu Y, Bieschke J, Gao J, Dendle MA, Kelly JW. Amide-to-E-Olefin versus Amide-to-Ester Backbone H-bond Perturbations: Evaluating the O-O Repulsion for Extracting H-Bond Energies. J. Am. Chem. Soc. 2006; 128:15948-15949.
- Gao J, Bosco DA, Powers ET, Kelly JW. Localized Thermodynamic Coupling between Hydrogen Bonding and Microenvironment Polarity Significantly Stabilizes Proteins.Nature Struct. Mol. Biol. 2009; 16:684-690.
- Jager M, Dendle M, Kelly JW. Sequence Determinants of Thermodynamic Stability in a WW domain–an all β-sheet protein. Protein Sci. 2009; 18:1806-1813.
- Fuller AA, Du D, Liu F, Davoren JE, Kroon G, Powers ET, Wipf P, Gruebele M, Kelly JW. Evaluating β-turn Mimics as β-sheet Folding Nucleators. Proc. Natl. Acad. Sci. 2009; 106:11067-11072
Introduction
Transthyretin amyloid diseases: understanding the mechanism of proteotoxicity and inhibition of amyloid fibril formation
Understanding Abeta aggregation and Alzheimer's disease
Gelsolin amyloid disease mechanism and therapeutics
β-Sheet folding
Parkinson's disease
Restoration of enzyme homeostasis in lysosomal storage diseases
Amyloid as a functional fold
California: 10550 North Torrey Pines Road, La Jolla, CA 92037 - (858) 784-1000
Florida: 130 Scripps Way, Jupiter, FL 33458 - (561) 228-2000
Copyright © , The Scripps Research Institute (TSRI). All Rights Reserved.