Publications
| Nucleosome stability safeguards cell identity, stress resilience and healthy aging Tan, E. P. et al. |
|
|




| Autophagy activator AA-20 improves proteostatis and extends Caenorhabditis elegans lifespan Tan, E. P. et al. |
|
|




| AI-Driven Identification of Exceptionally Efficacious Polypharmacological Compounds That Extend the Lifespan of Caenorhabditits elegans Avchaciov, K. et al. |
|
|


| Uncoupling overeating and fat storage by modulation of different serotongeric receptors Clark, C. et al. |
|
|




| TargetSeeker-MS: A Bayesian Inference Approach for Drug-Target Discovery Using Protein Fractionation Coupled to Mass Spectrometry Xie, Y. et al. |
|
|




| Activity-dependent synthesis of Emerin gates neuronal plasticity by regulating proteostasis Xie, Y. et al. |
|
|




| Quantifying Food Intake in Caenorhabditis elegans by Measuring Bacterial Clearance Clark, C. et al. |
|
|



| Proteostasis is differentially modulated by inhibition of translation initiation or elongation Clay, KJ. et al. |
|
|




| Targeting Clic1 for the treatment of obesity: A novel therapeutic strategy to reduce food intake and body weight Zapata, RC. et al. |
|
|




| Adipocytes control food intake and weight regain via Vacuolar-type H+ ATPase Zapata, RC. et al. |
|
|




| Rapamycin-mediated mouse lifespan extension: Late-life dosage regimes with sex-specific effects Strong, R. et al. |
|
|



| A chemical biology approach to identifying molecular pathways associated with aging Currais, A. et al. |
|
|




| The aging transcriptome: read between the lines Perez-Gomez, A. et al. |
|

| Elevating acetyl-CoA levels reduces aspects of brain aging Currais, A. et al. |
|
|




| Pharmacological convergence reveals a lipid pathway that regulates C. elegans lifespan Chen, A.L. et al. |
|
|




| Loss of genomic integrity induced by lysosphingolipid imbalance drives ageing in the heart Ahuga, G et al. |
|
|




| A phenotypic Caenorhabditis elegans screen identifies a selective suppressor of antipsychotic-induced hyperphagia Perez-Gomez, A et al. |
|
|




| Translation attenuation by minocycline enhances longevity and proteostasis in old post-stress-responsive organisms Solis, GM. et al. |
|
|




| Geroneuroprotectors: Effective Geroprotectors for the Brain Shubert, D. et al. |
|
|

| The mitochondrial ATP synthase is a shared drug target for aging and dementia Goldberg J. et al. |
|
|




| Computation Analysis of Lifespan Experiment Reproducibility Petrascheck M and Miller DL |
|
|




| The DrugAge database of aging-related drugs Barardo D, et al. |
|
|


| C. elegans as Model for Drug Discovery
Carretero M, Solis GM, Petrascheck M |
|
| Mood, stress and longevity: convergence on ANK3
Rangaraju S, Levey DF, Nho K, Jain N, Andrews KD, Le-Niculescu H, Salomon DR, Saykin AJ, Petrascheck M, Niculescu AB |
|
|







| Metabolic drift in the aging brain
Ivanisevic J, et al. |
|
|




| Proton Pump Inhibitors Accelerate Endothelial Senescence Yepuri G, Sukhovershin R, Nazari-Shafti TZ, Petrascheck M, Ghebre YT, Cooke JP |
|
|




| C. elegans S6K Mutants Require a Creatine-Kinase-like Effector for Lifespan Extension McQuary P, et al. |
|
|





| Suppression of transcriptional drift extends C. elegans lifespan by postponing the onset of mortality
Rangaraju S, Solis GM, Thompson R, Gomez-Amaro R, Kurian L, Encalada S, Niculescu III A, Salomon D, Petrascheck M |
|
|







| Atypical antidepressants extend lifespan of Caenorhabditis elegans by activation of a non-cell-autonomous stress response Rangaraju S, Solis GM, Andersson S, Gomez-Amaro R, Kardakaris R, Broaddus C, Niculescu III A, Petrascheck M |
|
|






| Measuring Food Intake and Nutrient Absorption in Caenorhabditis elegans
Gomez-Amaro R, Valentine E, Carretero M, LeBoeuf S, Rangaraju S, Broaddus C, Solis GM, Williamson J, Petrascheck M |
|
|





| Pharmacological classes that extend lifespan of Caenorhabditis elegans Carretero M, Gomez-Amaro R, Petrascheck M |
|
| High-throughput small-molecule screening in Caenorhabditis elegans Rangaraju S, Solis GM, Petrascheck M |
|
|


| The metabolite α-ketoglutarate extends lifespan by inhibiting ATP synthase and TOR Chin RM, et al. |
|
|



| A pharmacological network for lifespan extension in Caenorhabditis elegans Ye X, Linton JM, Schork NJ, Buck LB, Petrascheck M |
|
|





| Measuring Caenorhabditis elegans life span in 96 well microtiter plates
Solis GM, Petrascheck M |
|
|
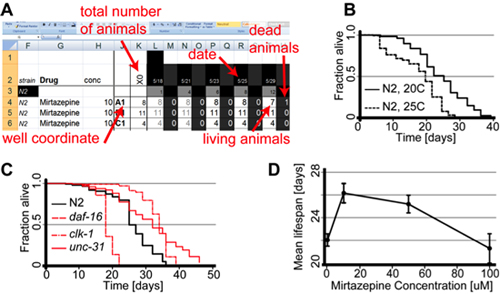
| A high-throughput screen for chemicals that increase the lifespan of Caenorhabditis elegans
Petrascheck M, Ye X, Buck LB |
|
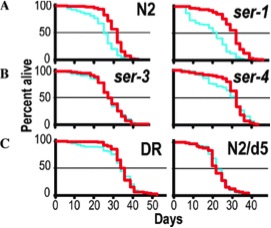
| An antidepressant that extends lifespan in adult Caenorhabditis elegans
Petrascheck M, Ye X, Buck LB |
|
|


