Ribozyme Reversal of Fortunes
By Jason Socrates
Bardi
"The
one duty we owe to history is to rewrite it."
—Quote
attributed to Oscar Wilde (1854–1900)
Ten years ago, a group of scientists who were all working
in the field of RNA processing met at the annual meeting of
the RNA Society in Madison, Wisconsin to discuss their latest
research.
Their field had gone through a lot in the previous decade.
RNA processing encompasses the biochemical mechanisms through
which RNA chains are edited—cut and pasted into new RNA
chains, much as individual shots in a movie are spliced together
by an editor to make a scene. And until the early 1980s, scientists
had assumed that only proteins spliced RNA. For that matter,
scientists had assumed that only proteins could carry out
any enzymatic reaction at all.
In 1982, however, Tom Cech at the University of Colorado
Boulder made the completely unexpected discovery that RNA
from a unicellular protozoan called Tetrahymena thermophila
could perform auto-splicing. Cech shared the Nobel Prize in
Chemistry in 1989 with Sidney Altman for his discovery of
RNA molecules with catalytic function, and this opened up
a whole new field for scientists: the structure, function,
and mechanism of action of RNA enzymes.
Scripps Research Institute Associate Professor Martha Fedor
was a postdoctoral fellow in the Boulder laboratory of Olke
Uhlenbeck in 1989, and was next door to Cech when the significance
of Cech's discovery was acknowledged by the Nobel committee.
She did some of the first quantitative measurements of the
kinetic parameters of RNA enzymes, which soon began to be
called "ribozymes."
Fedor, who is now a member of The Skaggs Institute for Chemical
Biology at Scripps Research, was still investigating RNA enzymes
10 years later when leading scientists in the ribozyme field
gathered for this 1994 meeting. The mood at the meeting was
one of growing confidence and elation. Dozens of ribozymes
had been characterized, and the way in which they catalyzed
reactions was known: ribozymes had to bind to metal cofactors
to function. In fact, some scientists were so confident of
this that they had taken to calling ribozymes "metalloenzymes."
But this confidence was misplaced.
Only a few years later, Fedor demonstrated that merely positively
charged ions were needed to enable ribozymes to assemble into
functional structures. Metal ions need not participate directly
in catalytic chemistry.
Fedor and her colleagues saw that ribozymes can react in
alternative buffers like cobalt hexamine or ammonium acetate
instead of conventional metal cations like magnesium or manganese.
Ammonium acetate buffers contained no metal ions at all. In
cobalt hexamine, the metal remains bound to its amine ligands
and cannot facilitate reactions by binding ligands associated
with the reactive phosphate in the way that magnesium or manganese
ions would. Cobalt hexamine does contain metal, but the metal
is complexed in such a way that it cannot facilitate reactions
the way that traditional metal cations would.
"Our claim to fame was that our ribozyme [functioned] perfectly
well in an environment with no [metal] cofactors," she says.
Now, says Fedor, a decade after that Madison meeting, ribozymes
have undergone a complete reversal of fortune in terms of
being characterized as metalloenzymes—in fact, now scientists
wonder whether any ribozymes use metal cofactors.
"Our field is a wonderful one," says Fedor. "People are
very good about saying, 'Everything we thought we knew was
wrong.'"
RNA Minimalism
The discovery of catalytic RNA in 1982 altered what had
been known as the central dogma of molecular biology—that
genetic information was stored in DNA molecules and that biological
action (catalysis) was taken by proteins.
RNA had largely been assumed to be an intermediary in this
process, unable to participate directly in catalysis.
Perhaps for good reason.
Protein enzymes are polymer strings made up of the 20 different
amino acid building blocks, many of which have reactive side
chains that can participate in biocatalysis. RNA polymers
are made up of only four different building blocks, the RNA
nucleotides, which lack the chemical versatility and reactivity
of the protein functional groups.
After the discovery that RNA could act as a biocatalyst,
many scientists began to theorize and discuss the possibility
of an RNA world—the notion that at one time the world
was ruled by RNA-based life forms in which RNA enzymes were
the chief catalytic molecules and RNA nucleotides were the
building blocks that stored genetic information. This raised
a number of questions, such as where catalysis came from and
how the world transitioned from an RNA-based form to the current
DNA-RNA-protein form.
Another group of scientists, including Fedor, became interested
in the fundamental question of how. How do ribozymes do it?
What is it about RNA structure that confers unique properties
on the nucleotides that allows them to do catalytic chemistry?
Today, half of Fedor's group at Scripps Research works on
understanding the mechanisms of RNA catalysis using a simple
ribozyme as a model—a shortened, "minimal" form of catalytic
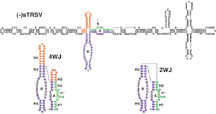
RNA that's about 50 nucleotides long.
Called the hairpin ribozyme, this minimal RNA enzyme was
discovered by botanists in plants infected by tobacco ring
spot virus and a number of other viruses. It is what is known
as plant satellite RNA.
Satellite RNAs are parasitic pieces of RNA that are not
exactly viruses because they don't encode for proteins. Instead,
they catalyze simple cut-and-paste reactions in order to replicate
themselves, exacerbating or ameliorating diseases caused by
plant viruses. A plant that has tobacco ring spot virus, for
instance, will be more diseased if it also has this satellite
RNA.
Since the hairpin ribozyme is so short, it's easily handled
in the laboratory, which allows Fedor and her colleagues to
manipulate individual parts of the ribozyme to see which ones
are most important for catalysis.
When the high-resolution structure of the hairpin ribozyme
appeared a few years ago, determining which parts are important
for catalysis became easier because the structure pointed
to particular bases that might have a role in the catalysis
by virtue of their proximity to the ribozyme's active site.
By eliminating these bases one by one within the active
site of the ribozyme, Fedor and her colleagues were able to
determine which ones were important for catalysis. Eliminating
some nucleotides resulted in no change in catalysis, suggesting
that these RNA bases played no significant role. Eliminating
others resulted in hairpin ribozymes with defects, suggesting
that these bases contribute to normal activity.
The scientists have concentrated on these latter nucleotides,
hoping to learn something about the ribozyme catalytic mechanism
through a technique called exogenous nucleobase rescue of
abasic substitutions—a way they found to delete particular
nucleobases from the active site of an RNA enzyme without
disrupting its structure and restore activity by providing
nucleobases in solution.
"It's a cool way of investigating the structural and functional
requirements for a particular nucleotide," says Fedor, adding
that the technique was a big breakthrough. "Before we found
that abasic substitutions could be rescued by exogenous nucleobases,
any time we touched [the ribozyme], we destroyed the structure
and couldn't tell what was important for holding it together
and what was important for catalytic chemistry."
Exogenous nucleobase rescue involves substituting RNA nucleotides
of interest with abasic residues that maintain the continuity
of the RNA chain but have a single hydrogen atom in place
of a critical nucleobase. Then the researchers add in small
molecules that can rescue the activity that was lost due to
the missing nucleobase.
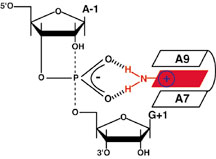
Using this technique, Fedor has been developing a new picture
of the RNA enzyme reaction mechanism.
She is proposing that rescued nucleobase ribozymes bind
the reactive phosphate through the same functional groups
that normally engage in Watson-Crick base pairing in conventional
RNA structures. In the context of the ribozyme active site,
however, these interactions promote catalysis by stabilizing
a highly energetic transition state in which five oxygen atoms
are associated with one central phosphorous atom—a highly
electronegative situation. A good way to stabilize this transition
state would be to have a positive charge close by, a charge
that might come from adding a proton to a nearby nuclotide.
Data showing that rescue activity increases as reaction buffers
are made increasingly acidic supports this idea, she says.
The way in which RNA functions as an enzyme is of even more
interest now that the ribosome structure has been solved.
The ribosome, a huge nucleoprotein complex, is the RNA machine
that strings together amino acids to make proteins in cells,
and its structure reveals that there are no proteins or metal
ions in its active site. It is the RNA alone that performs
the peptide bond formation.
"This is one of the most well-studied reactions on the planet,
but we still don't know how the reaction is catalyzed," says
Fedor.
In Vivo Approaches and Therapy
The other half of Fedor's group uses the ribozyme as a model
for RNA folding in vivo. Using yeast as a model system, they
look at ribozyme activity and ask how their experiments carried
out in vitro will translate to living systems.
Interest in ribozyme activity in vivo has grown in
the last few decades because of its potential for use in therapeutic
applications to human diseases. There are many diseases that
are associated with aberrant RNA expression that could be
addressed by modifying RNA levels—from knocking out mRNAs
expressed by cancerous oncogenes to getting rid of viral RNA
inside infected cells.
"Ribozymes that recognize targets through complementary
base pairing allow you to develop a therapeutic reagent without
knowing anything about the structure or the function of your
protein target—you just need to know the gene," says
Fedor.
Fedor and her group have developed a way of measuring ribozyme
cleavage of RNA targets in vivo, based on the intrinsic
rate of RNA degradation. By correlating the rate at which
the RNA disappears in the presence or absence of the ribozymes,
they can determine the cleavage rate of the ribozymes.
"All RNAs turn over at a characteristic rate," says Fedor.
"We use this as a clock."
This is important in the context of therapeutic RNA because
the cleavage rate determines the potential efficacy of the
RNA. Only if the cleavage rate is much greater than the intrinsic
degradation rate will the RNA catalysis be worthwhile. If
the cleavage rate is not much greater than the intrinsic degradation
rate, then subjecting cells with it probably will not make
a difference in the abundance of the target.
Inactivation of target RNA is most effective against a stable,
slowly decaying RNA, such as a chronic viral infection like
hepatitis where the RNA persists for a long time in cells.
On the other hand, the stability of the ribozyme structure
is a factor as well, and Fedor has done experiments with the
natural, long form of ribozyme to compare its stability to
the short form.
"One of the things we've learned over the last few years
that has made a big difference is that the natural form assembles
in a four-way helical junction."
This four-way helical junction is far more stable than the
two-way helical junction of the short hairpin ribozyme and
is completely functional at intracellular ionic strength,
whereas the minimal, two-way junction ribozymes require high
concentrations of divalent cations for its activity.
Fedor's results suggest that four-way junction ribozymes
may be more amenable to therapeutic applications because the
four-way junction ribozymes are more stable in vivo.
Data from Fedor lab studies also show that the four-way
junction ribozymes are very efficient ligases—that is,
they can stitch nucleotides together as well as they can pull
them apart. This suggests that the four-way junction ribozymes
could be used as RNA repair enzymes.
Such findings may help the therapeutic dreams of ribozyme
enthusiasts become a reality.
Send comments to: jasonb@scripps.edu

|