Picking Up the Pieces
By Jason Socrates
Bardi
"Presently
I am going to press the lever, and off the machine will
go... Have a good look at the thing."
—H.G.
Wells, The Time Machine, 1895.
Professor Jamie Williamson, who is a member of the Department
of Molecular Biology and The Skaggs Institute for Chemical
Biology at The Scripps Research Institute, remembers one day
six years ago as if it were yesterday—the day that the
ribosome stopped being just a big purple blob.
A big purple blob was how anyone who had studied molecular
biology from textbooks published in the mid-1990s knew the
shape of the ribosome, the molecular "engine" that drives
protein synthesis and is fundamental to the life of all cells
in nature. That crude shape was based on electron micrograph
images from the 1970s.
But, at the 1998 the triennial ribosome conference meeting
in Denmark, several groups presented fairly high-resolution
ribosomal structures. On that day, the larger scientific world
got its first close-up look at the ribosome.
"It was totally clear that this was a major advance in structural
biology," says Williamson. "It's a rare moment in your career
when you are right in the middle of something that you know
is going to [eventually] win the Nobel Prize."
Not everything seemed so joyous at the time, though.
"It was a time of horror and wonder," admits Williamson,
who had been working on the structures of small pieces of
the ribosome, aiming to piece them together into a complete
picture of the molecule once he was done. "I had the dubious
honor of reviewing all the papers, which meant I was qualified
to review them but was not publishing any of them. But after
awhile, I sobered up and realized that [the new structures]
opened up many doors for new kinds of experiments."
It has been a wild ride ever since.
Two Easy Pieces
High-resolution structures of molecules like the ribosome
are as important to biologists as detailed blueprints are
to mechanics. And just as an auto mechanic needs to have an
accurate picture of a car engine to understand how it drives
the car, so do biologists need detailed blueprints of the
ribosome to understand how it drives protein synthesis.
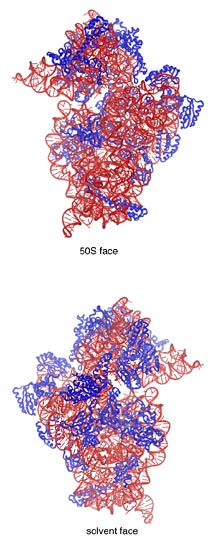
One of the ancient structures in biology, the ribosome is
a huge molecule composed of two subunits, one slightly larger
than the other. These are called the 50S and 30S subunits
in bacterial ribosomes and the 60S and 40S subunits in mammalian
ribosomes—names that date back decades to early biochemical
experiments. (The "S" stands for Svedberg, a measure of the
sedimentation of a molecule in a centrifuge).
Williamson has been interested in RNA–protein interactions
since the early years in his career, when he solved RNA–protein
complexes with his laboratory team using nuclear magnetic
resonance (NMR). Because of this, the ribosome—which
is one big mess of protein and RNA interactions—intrigued
him.
The ribosome molecule is a complex of about two-thirds RNA
and one-third protein by mass. In bacteria, the large 50S
subunit is an amalgam of two RNA chains and 31 different proteins.
For the most part, these ribosomes are RNA machines. At their
core is the RNA, which does all the important work, holding
the two subunits together and recognizing the mRNA, for instance.
The proteins are, for the most part, on the outside and contribute
to the structural stability of the ribosome as well as recruiting
other molecules that are needed for translation.
The 50S subunit is often referred to as the catalytic subunit
because it contains the peptidyl transferase center—the
part of the molecule that does the chemistry of joining two
amino acids together in a growing peptide chain.
The smaller 30S subunit has a single RNA chain and 21 different
proteins. Its job is to read the genetic code in messenger
RNA (mRNA) and pair the nucleotide codons with their corresponding
amino acids.
Studying Ribosome Assembly One Piece at a Time
To study the structure, Williamson and his laboratory divided
up the 30S subunit of the ribosome piece-by-piece and sought
to solve each piece at high resolution and then stitch the
structures of all the pieces together into the low-resolution
electron microscope image. Now Williamson is asking different
questions, such as how the ribosome is assembled.
In fact, in the few years since the ribosome morphed from
a purple blob to a detailed atomic structure, the field has
only become more exciting as many scientists have been posing
more interesting questions.
Scientists are also further refining their ribosomal structures
to get clearer and clearer pictures of it. They can do things
not possible a few years ago, such as make crystals of the
ribosome, soak it in drugs known to bind to it, and find where
these drugs bind. And scientists are solving the structures
of ribosomes from mammals and other eukaryotic organisms to
compare these to the bacterial ribosomes.
Now that there are structures available for the ribosome,
says Williamson, scientists are starting to look at how the
structure of the molecule contributes to its function and
are reinterpreting decades of biological experiments using
the structure. Many questions are still unanswered. It is
not exactly clear how the 50S subunit catalyzes peptide bond
formation, for instance, nor how the ribosome slides down
the mRNA as it translates.
"In the end," he says, "we are doing better experiments
now than we ever did before."
Part of his laboratory focuses on the assembly process of
the 30S subunit, asking questions focusing on the initial
events that cause its 21 proteins to come together with its
1541-base RNA chain, the order of the pieces coming together,
and the conformation changes occurring in the process.
He is able to do this because the strategy that he and his
colleagues used to make the small pieces of the ribosome for
their structural studies was based on an assembly "map" of
the ribosome that was created in biochemical studies in the
1960s an 1970s. The scientists who did these studies took
the ribosome apart, purifying all its pieces separately and
then reconstituting them one-by-one, tinkering with the order
in which they mixed the components back together. Because
the ribosome self-assembles, by tinkering with this order
the scientists were able to discover the steps in which the
ribosome assembles.
"What we did [based on this map]," says Williamson, "was
to build up a whole series of ever-bigger particles that corresponded
to the way in which this thing assembles."
After he saw no need to continue with his structural studies,
Williamson realized that he could use the pieces he was working
on for folding studies—looking at how pieces twist into
the final active conformation and providing a mechanistic
framework of how it gets there.
To do this, members of the Williamson laboratory use all
the tools of biochemistry, including fluorescence, calorimetry,
and mass spectrometry, to observe when the RNA of the 30S
subunit folds and when the proteins that are part of it hop
on.
"What we're trying to think about is how we can make a conceptual
film of the process, and we're trying to understand the fundamental
principles that embody what happens," says Williamson. "We
can all see the engine under the hood, and now we're trying
to understand how it is assembled."
New Targets in HIV
Another part of Williamson's laboratory focuses on the design
of new drugs to combat human immunodeficiency virus (HIV).
HIV, the virus that causes AIDS, has a small genome of around
10,000 nucleotides, so it only codes for a handful of proteins.
Some of these have familiar-sounding names, like the protease
or the reverse transcriptase, because there are already drugs
on the market that target them.
Some of the other proteins HIV encodes, like the regulatory
proteins Tat and Rev, have not had drugs designed to block
them, despite the fact that they are important in the viral
lifecycle. Rev, for instance, is a nuclear transport factor
that binds to HIV mRNA and is required for getting the full-length
HIV RNA out of the nucleus so that it can be translated into
the long envelope and capsid proteins and packaged into new
virions.
Williamson and his laboratory solved the first structure
of Rev bound to RNA in 1996.
Now in a new grant titled "Structural Biology for AIDS-Related
Targets" that was funded by the National Institutes of Health
(NIH) beginning on October 1 of last year, he and several
colleagues at Scripps Research are looking for new or underexploited
targets against which to develop novel drugs. In particular,
they are attempting to learn how to target the HIV RNA.
"Nobody has really done drug design or drug discovery against
an RNA target," says Williamson.
One of the targets that Williamson is looking at is a section
of the viral RNA to which the HIV protein Rev binds called
the Rev response element (RRE), RRE is a 350 nucleotide section
of the viral mRNA transcript that folds up into a stable binding
site for Rev. Binding of Rev to RRE is a key stage in viral
replication. Significantly, this region seems to be completely
conserved—unlike the rest of the HIV genome, it is not
prone to mutate.
"In principle," says Williamson, "if you could block this
interaction, you could inhibit viral replication."
At the moment, Williamson is collaborating with several
other investigators at Scripps Research funded through this
grant. These include Assistant Professor Mirko Hennig, who
is working with Williamson to set up NMR screens to assay
the ability of small molecules to block the interaction between
Rev and RRE; Associate Professor Davis Millar, who has a fluorescent
screen that can also assay the binding of Rev to RRE; Professor
Larry Gerace, who has assays set up in cells to look for Rev
function; and Associate Professor Hartmuth Kolb, a chemist
who has his own library of compounds and the ability to make
analogues of any promising leads they find.
Williamson notes, "All the stuff we're doing [targeting
HIV RNA] can be applied to the ribosome to develop new antibiotics."
Full Circle
Antibiotics are the notable exception to the rule that most
drugs target proteins. Drugs like tertracycline, kanamycin,
and erythromycin all work by targeting bacterial ribosomes.
They bind to different parts of the ribosome and interfere
with its ability to translate mRNA into protein.
It's not clear how all of these antibiotics do this, but
some evidence suggests that certain antibiotics lead to the
disassembly of ribosomes. One intriguing possibility is that
scientists could develop antibiotics that target bacterial
ribosomes specifically and interfere with their assembly,
"essentially throwing sand into the gear box," says Williamson.
In one corner of Williamson's office is an old foam mouse
pad adorned with the ribosome, a freebie produced to celebrate
and promote the long-awaited publication of a high-resolution
structure. It is not the purple blob of old but the atomic
shape of post-1998.
Still, it sits unused on a corner table, the victim of better
mouse technologies that use laser motion detectors instead
of track balls.
There is still a long way to go with the ribosome, says
Williamson.
Send comments to: jasonb@scripps.edu

|