|
(page 2 of 2)
Cell Crawling and Wound Healing
Understanding the signals and interactions driving the molecules
that move the cells should help to elucidate the mechanisms
that are common to a number of specific health-related problems
studied in the Laboratory for Cell Motility.
Any time the skin is cut, the body will work to heal the
wound. Wounds will over time close and the skin will grow
back, connecting the two sides of the cut.
What drives this process is the action of the cells, which
crawl forward to close and heal the wound. Once the two sides
meet, the cells stop crawling and simply adhere to each other
to make a solid tissue again, in a process known as contact
inhibition of cell motility.
One area that interests Waterman-Storer is how the cells
stop crawling once they contact each other. Presumably when
the leading edge contacts another cell, a signal cascade occurs
that ultimately shuts down the polymerization of the actin
and halts cell motility.
"But what is happening to the cytoskeleton when that happens?"
she asks. "What is regulating it and what are the kinetics
when that occurs?"
Another area of interest is that of embryonic development.
Cell motility is of huge importance to this area, because
stem cells that will develop into nerve cells have to move
to the correct location in the embryo before they can form
nerve tissue and extend their axons so they can communicate
with other cells.
The problem of motility is also closely related to cancer
studies, since metastasis of cancerous cells is caused by
the loss of this contact inhibition in cell motility. In fact,
metastatic cells can be identified on plates by their ability
to form colonies that continue to crawl and divide.
"If there were a way to selectively control tumor cell motility,"
says Waterman-Storer, "It could be used as an anti-metastatic
therapeutic agent." Then cancers would remain local, forming
tumors that could be easily excised.
And in the Laboratory for Cell Motility, this understanding
starts with microscopy of the live cells in migration.
Capturing Movement
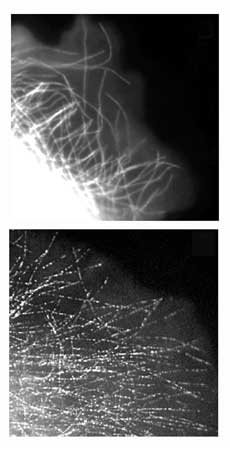
Waterman-Storer uses a spinning disk confocal microscope
with excellent optics to produce the images with low background
florescence. Images are captured with a high-resolution charge
coupled device (CCD), which was originally designed for astronomers
and is electronically cooled to improve the signal-to-noise.
"These have improved greatly over the last five years," says
Waterman-Storer.
The cameras have gotten so good, in fact, that they can
now image cells at the optical limit of diffraction, capturing
images with a rich dynamic range where each pixel represents
an area of about six microns. And the cameras are so sensitive
that they can detect groups of only two or three fluorescently
labeled molecules above the noise.
To image these cells over time, Waterman-Storer captures
a frame every 10 seconds and uses a mechanical filter wheel
to take nearly simultaneous exposures at several wavelengths
in order to capture the overall dynamics on the cell. One
image will be of microtubules that fluoresce at one wavelength,
and another will be of the actin, which fluoresce at another.
"One of the specialties of the laboratory is multimode imaging,"
says Waterman-Storer, "the ability to look at multiple probes
simultaneously."
Multimodal imaging can be used to look at whether the microtubules
track along the actin bundles or are pushed in the same direction
as actin moves backwards in the cell. Viewing this sort of
relationship is evidence that the two cytoskeletal structures
contact each other and stick together.
Then this data can be correlated with in vitro assays that
characterize which particular proteins are mediating these
interactions.
Between Two Worlds
The interaction of different types of cytoskeletal proteins
puts Waterman-Storer between two worlds, so to speak.
These cytoskeletal proteins are polar, and cells use their
polarity to generate polarized cell morphology. Everything
from the formation of a microtubule spindle for mitotic cell
division to adhesion and movement of the cells comes from
this polarity.
And just as these proteins polarize the cells so have they
polarized cell biologists, who have traditionally fallen into
separate camps dedicated to studying the distinct proteins,
either microtubules or actin.
Actin research has benefited from years of biochemical studies
of the molecule and the myriad proteins that bind to and regulate
it. But while many of these regulatory molecules have been
characterized biochemically, nobody has ever looked at their
effect directly in cells. FSM has changed that completely.
"FSM has provided a way to quantitatively analyze actin
dynamics in vivo," says Waterman-Storer. "And you can study
all the actin in a cell at one time."
Actin filaments form bundles and cross-linked meshes, making
it difficult to see individual filaments with traditional
florescence. Another technique, photobleaching, had previously
been used, but it was time-consuming and technically demanding.
Also, this technique was difficult to use for studying actin
in the whole cell since only a small part of the cell could
be photobleached.
Microtubules, for their part, have had a long history under
the microscope. They have the ability to rotate polarized
light, so they have been studied in vivo for many years. But
what people haven’t done is to look at how proteins that
bind to actin affect microtubule dynamics and vice versa.
"There are some old observations that tell us their interactions
are important," says Waterman-Storer. "But nobody has really
followed up to figure out the kinetics of these interactions,
what mediates them, and how they are regulated."
Years ago, a Russian biologist observed that when microtubules
are destroyed in a cell, the actin depolarizes and cell motility
stops, leading many to believe that microtubules provide instructions
to the actin in a cell. Microtubules are known to bind regulator
molecules of the actin-mediating GTPase signaling molecule,
but the mechanism is not yet clear.
"What is clear," says Waterman-Storer, "is that we can image
them simultaneously to look for evidence if interactions between
the two."
Some little motion in the cell catches my eye and Waterman-Storer
follows my gaze to the computer screen. She is in mid-sentence,
saying, "Every genesis involves huge migrations of cells.
You have these neural crest cells..." and her words trail
off.
"I love watching these movies." She says.
1 | 2 |

|