The Wittenberg Laboratory
Research
Control of
Cell Cycle Initiation. 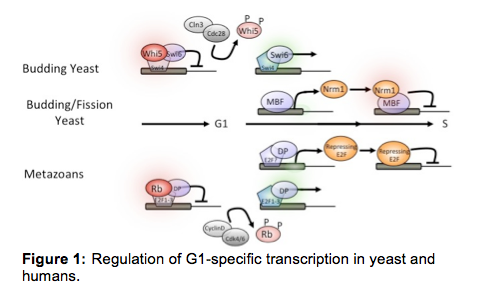 Progress through the
eukaryotic cell cycle is driven by sequential waves of accumulation of regulatory
proteins that determine the sequence of cell cycle events (Wittenberg and Reed,
2005). As a consequence, periodic transcription regulates the sequence and
timing of cell cycle events and it is, in turn, regulated by those events. Our laboratory studies the regulation
of G1/S transcription (sometimes called G1-specific transcription), the wave of
expression associated with commitment to a new cell cycle. The G1/S gene family
in the budding yeast is comprised of about three hundred genes that are
co-expressed as cells initiate a new cycle during G1 phase. Those genes are
expressed under the control of two distinct transcription factors known as SBF
and MBF (Fig. 1). We have identified two transcriptional repressors that
regulate the G1/S transcription factors. Whi5 specifically associates with and
represses SBF-dependent transcription during early G1 phase (de Bruin et al.,
2004). It is inactivated as a consequence of phosphorylation by the G1
cyclin/CDK complex, Cln3/Cdc28 leading to SBF activation. The second repressor,
Nrm1, acts as a corepressor together with MBF to repress transcription as cells
exit G1 phase (de Bruin et al., 2006). Nrm1 is encoded by an MBF target and,
thereby, participates in a negative feedback loop that limits G1-specific
transcription. Furthermore, Nrm1
is antagonized by the Rad53 checkpoint kinase in cells responding to DNA
replication stress (Travesa et al, 2012). Whi5 and Nrm1 share a small domain
called the GTB motif that mediates their repressive interaction with their
cognate transcription factor (Travesa et al, 2013). Together, they impose the transcriptional repression that is central to the architecture of G1/S
regulatory circuitry. A similar wave of gene expression encoding many
orthologous proteins occurs during G1 phase in metazoans, including humans, and
is controlled by the E2F family of transcription factors. Deregulation of the Rb/E2F transcriptional
circuit is tightly associated with disease including cancer.
Progress through the
eukaryotic cell cycle is driven by sequential waves of accumulation of regulatory
proteins that determine the sequence of cell cycle events (Wittenberg and Reed,
2005). As a consequence, periodic transcription regulates the sequence and
timing of cell cycle events and it is, in turn, regulated by those events. Our laboratory studies the regulation
of G1/S transcription (sometimes called G1-specific transcription), the wave of
expression associated with commitment to a new cell cycle. The G1/S gene family
in the budding yeast is comprised of about three hundred genes that are
co-expressed as cells initiate a new cycle during G1 phase. Those genes are
expressed under the control of two distinct transcription factors known as SBF
and MBF (Fig. 1). We have identified two transcriptional repressors that
regulate the G1/S transcription factors. Whi5 specifically associates with and
represses SBF-dependent transcription during early G1 phase (de Bruin et al.,
2004). It is inactivated as a consequence of phosphorylation by the G1
cyclin/CDK complex, Cln3/Cdc28 leading to SBF activation. The second repressor,
Nrm1, acts as a corepressor together with MBF to repress transcription as cells
exit G1 phase (de Bruin et al., 2006). Nrm1 is encoded by an MBF target and,
thereby, participates in a negative feedback loop that limits G1-specific
transcription. Furthermore, Nrm1
is antagonized by the Rad53 checkpoint kinase in cells responding to DNA
replication stress (Travesa et al, 2012). Whi5 and Nrm1 share a small domain
called the GTB motif that mediates their repressive interaction with their
cognate transcription factor (Travesa et al, 2013). Together, they impose the transcriptional repression that is central to the architecture of G1/S
regulatory circuitry. A similar wave of gene expression encoding many
orthologous proteins occurs during G1 phase in metazoans, including humans, and
is controlled by the E2F family of transcription factors. Deregulation of the Rb/E2F transcriptional
circuit is tightly associated with disease including cancer.
Role of the SCFGrr1
E3 Ubiquitin Ligase in Nutrient Uptake and Cell Proliferation. The
role of ubiquitylation is pervasive in biological regulatory mechanisms and
defects in that process are associated with numerous human diseases. Ubiquitin-mediated proteolysis, as well
as non-proteolytic processes involving ubiquitylation, is mediated by an
enzymatic cascade that, like ubiquitin itself, is highly conserved throughout
eukaryotes. One of the primary determinants of specificity of ubiquitylation
occurs at the level of substrate selection. That process is mediated, in most cases, by the E3 component
of the cascade, the ubiquitin ligase. Because of the diversity of proteins
regulated by ubiquitylation the family of ubiquitin ligases is large and
diverse.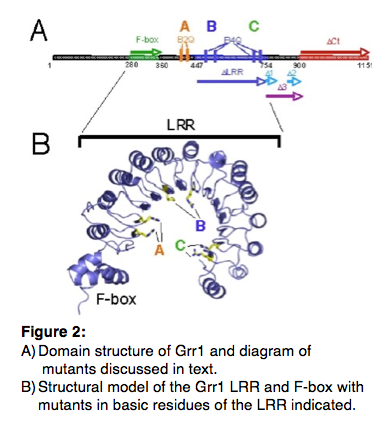 #160; Among E3 ligases, the SCF
(Skp1-Cullin-F-box protein) family is the largest. The diversity is primarily a
consequence of the diversity of F-box proteins. Our laboratory has studied the SCFGrr1
E3 ubiquitin ligase, a critical element of both nutritional regulation of cell
growth (Spielewoy et al, 2004; 2010) and cell cycle control (Lanker et al,
1995; Hsiung et al, 2001) targeting both transcriptional regulators and cyclin
proteins for ubiquitylation and degradation.
We have undertaken a detailed investigation of the nature of substrate
selection and discrimination by the SCFGrr1 E3 ubiquitin ligase
(Lanker et al, 1995; Hsiung et al, 2001; Berset et al, 2002; Spielewoy et al,
2004). The F-box Grr1 mediates the interaction with its substrates, in
part, via its leucine-rich repeat (LRR; Fig. 2). Because of its central role in
pathways critical for the regulation of cell growth and the diversity of
targets that it recognizes, establishing the basis for specificity of Grr1 is
expected to be particularly useful in developing an understanding of the manner
in which F-box proteins participate in biological regulatory mechanisms. Elucidation of the mechanisms by which
E3 ubiquitin ligases, in general, and LRR-containing F-box proteins, specifically,
recognize and regulate protein stability is of critical importance for the
development of treatments via drug targeting and other approaches.
#160; Among E3 ligases, the SCF
(Skp1-Cullin-F-box protein) family is the largest. The diversity is primarily a
consequence of the diversity of F-box proteins. Our laboratory has studied the SCFGrr1
E3 ubiquitin ligase, a critical element of both nutritional regulation of cell
growth (Spielewoy et al, 2004; 2010) and cell cycle control (Lanker et al,
1995; Hsiung et al, 2001) targeting both transcriptional regulators and cyclin
proteins for ubiquitylation and degradation.
We have undertaken a detailed investigation of the nature of substrate
selection and discrimination by the SCFGrr1 E3 ubiquitin ligase
(Lanker et al, 1995; Hsiung et al, 2001; Berset et al, 2002; Spielewoy et al,
2004). The F-box Grr1 mediates the interaction with its substrates, in
part, via its leucine-rich repeat (LRR; Fig. 2). Because of its central role in
pathways critical for the regulation of cell growth and the diversity of
targets that it recognizes, establishing the basis for specificity of Grr1 is
expected to be particularly useful in developing an understanding of the manner
in which F-box proteins participate in biological regulatory mechanisms. Elucidation of the mechanisms by which
E3 ubiquitin ligases, in general, and LRR-containing F-box proteins, specifically,
recognize and regulate protein stability is of critical importance for the
development of treatments via drug targeting and other approaches.
Ribosome Biogenesis in
Yeast (In
collaboration with Jamie Williamson, TSRI). The eukaryotic
ribosome biogenesis pathway involves the coordinated high level expression ~100
ribosomal protein (RP) genes, hundreds of ribosome biogenesis factor (RiBi)
genes, as well as the multicopy ribosomal RNA (rRNA) genes. While a great deal
is known about the inventory of RiBis, and the role of several of these in
specific steps in biogenesis, there is a tremendous amount that remains to be
discovered about the mechanism of binding of ribosomal proteins, and about the
specific roles of RiBis. We have recently initiated studies in collaboration
with the laboratory of Jamie Williamson at TSRI to provide a better
understanding of pathways for ribosomal protein association during biogenesis
and the connection of those pathways to cell growth and proliferation. 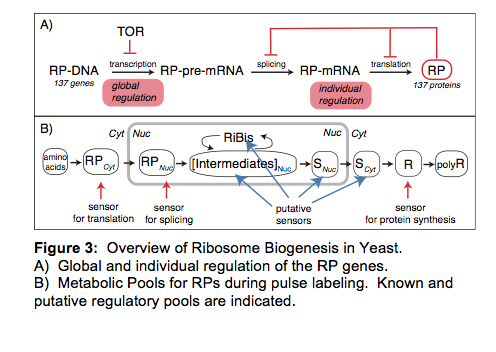 One of our main approaches is to
utilize stable isotope pulse labeling and quantitative mass spectrometry (QMS)
to probe the dynamics of RPs and intermediates in ribosome biogenesis. QMS
provides the opportunity to identify the protein composition in each of the
intermediate pools, and the rate of synthesis can be derived from the rate of
labeling (Fig.3).
Information about the role of RPs and RiBis can be obtained by
perturbing the pathway, and observing the resulting changes in the metabolic
pools. Cell growth is intimately related to the
biogenesis of the protein synthetic machinery. The impact of defects in
ribosome biogenesis factors on cell growth and cell cycle is well documented.
However, a systematic approach to understanding these defects has been lacking.
We propose to exploit our knowledge of
the pathways for ribosome biogenesis to interrogate the relationship between
that process, cell growth and cell cycle.
One of our main approaches is to
utilize stable isotope pulse labeling and quantitative mass spectrometry (QMS)
to probe the dynamics of RPs and intermediates in ribosome biogenesis. QMS
provides the opportunity to identify the protein composition in each of the
intermediate pools, and the rate of synthesis can be derived from the rate of
labeling (Fig.3).
Information about the role of RPs and RiBis can be obtained by
perturbing the pathway, and observing the resulting changes in the metabolic
pools. Cell growth is intimately related to the
biogenesis of the protein synthetic machinery. The impact of defects in
ribosome biogenesis factors on cell growth and cell cycle is well documented.
However, a systematic approach to understanding these defects has been lacking.
We propose to exploit our knowledge of
the pathways for ribosome biogenesis to interrogate the relationship between
that process, cell growth and cell cycle.
Selected
Publications:
Travesa, A., Kalashnikova, T.I., de Bruin, R.A.M., Cass, S.R., Chahwan, C., Lee, D.E., Lowndes, N., and Wittenberg, C. (2013). Repression of G1/S transcription is mediated via interaction of the GTB motif of Nrm1 and Whi5 with Swi6. Mol. Cell. Biol. In press.
Travesa, A., Kuo, D., de Bruin, R.A.M., Kalashnikova, T.I., Guaderrama, M., Thai, K., Aslanian, A., Smolka, M.B., Yates, J.R., Ideker, T., and Wittenberg, C. (2012). DNA replication stress differentially regulates G1/S genes via Rad53-dependent inactivation of Nrm1. EMBO J 31, 1811–1822.
Spielewoy, N., Guaderrama, M., Wohlschlegel, J.A., Ashe, M., Yates, J.R., and C. Wittenberg. (2010). Npr2, yeast homolog of the human tumor suppressor NPRL2, is a target of Grr1 required for adaptation to growth under suboptimal nutrient conditions. Eukaryotic Cell 9, 592–601.
de Bruin, R., Kalashnikova, T.I., Aslanian, A., Wohlschlegel, J.A., Chahwan, C., Yates, J., 3rd, Russell, P., and Wittenberg, C. (2008). DNA replication checkpoint promotes G1/S-specific transcription via Cds1-dependent phosphorylation of the MBF-bound repressor, Nrm1. Proc. Natl. Acad. Sci. USA 105, 11230-11235.
de Bruin, R., Kalashnikova, T.I., Chahwan, C., McDonald, W.H., Wohlschlegel, J.A., Yates, J., 3rd, Russell, P., and Wittenberg, C. (2006). Constraining G1-specific transcription to late G1-phase: The MBF-associated corepressor Nrm1 acts via negative feedback. Molecular Cell 23, 483-496.
Wittenberg, C., and Reed, S.I. (2005). Cell cycle-dependent transcription in yeast: promoters, transcription factors, and transcriptomes. Oncogene 24, 2746–2755.
Spielewoy, N., Flick, K., Kalashnikova, T.I., Walker, J.R., and Wittenberg, C. (2004). Regulation and recognition of SCFGrr1 targets in the glucose and amino acid signaling pathways. Mol. Cell. Biol. 24, 8994–9005.
de Bruin, R., McDonald, W.H., Kalashnikova, T.I., Yates, J., 3rd, and Wittenberg, C. (2004). Cln3 activates G1-specific Transcription via phosphorylation of the SBF-Bound Repressor, Whi5. Cell 117, 887-898.
Berset, C., Griac, P., Tempel, R., La Rue, J., Wittenberg, C., and Lanker, S. (2002). Transferable domain in the G(1) cyclin Cln2 sufficient to switch degradation of Sic1 from the E3 ubiquitin ligase SCF(Cdc4) to SCF(Grr1). Mol Cell Biol 22, 4463–4476.
Hsiung, Y.G., Chang, H.C., Pellequer, J.L., La Valle, R., Lanker, S., and Wittenberg, C. (2001). F-box protein Grr1 interacts with phosphorylated targets via the cationic surface of its leucine-rich repeat. Mol Cell Biol 21, 2506–2520.
Lanker,
S., Valdivieso, M.H., and Wittenberg, C. (1996). Rapid Degradation of the G1
Cyclin Cln2 Induced by CDK-Dependent Phosphorylation. Science 271,
1597–1601.
California: 10550 North Torrey Pines Road, La Jolla, CA 92037 - (858) 784-1000
Florida: 130 Scripps Way, Jupiter, FL 33458 - (561) 228-2000
Copyright © 2024, The Scripps Research Institute (TSRI). All Rights Reserved.