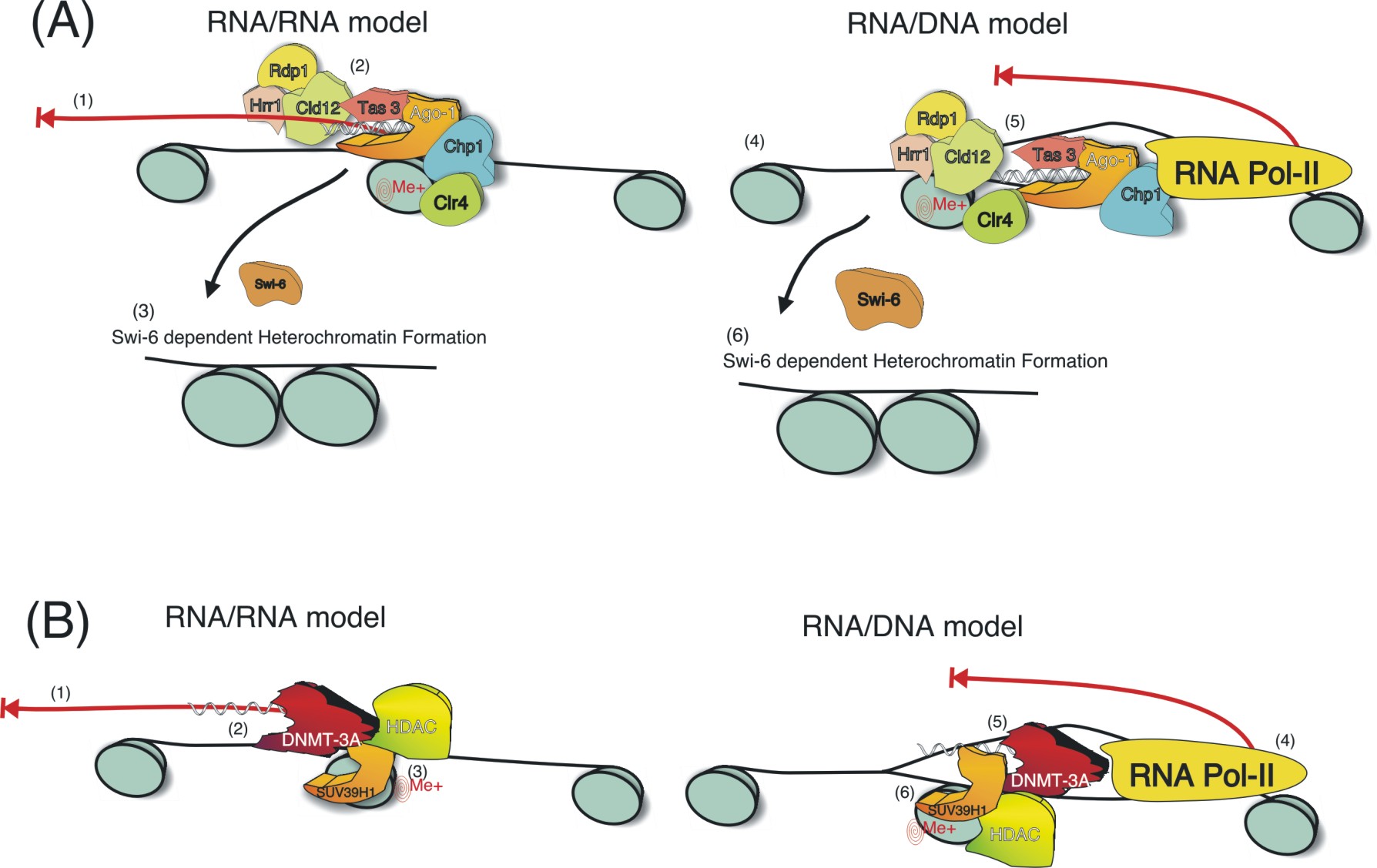
Proposed
Mechanisms for TGS in S. Pombe vs. Humans. Essentially two
models for siRNA mediated TGS are proposed here, either an RNA/RNA or a
RNA/DNA mediated mode of silencing. In S. Pombe (A) the siRNAs may
interact with a long non-coding transcript which spans the targeted
chromatin (1) subsequently allowing the RITS/RdRP complex to
localize to the targeted region (2) resulting in gene silencing
(3)(proposed by R. Allshire, personal communication). Alternatively,
siRNA mediated silencing may function through an RNA/DNA intermediate.
The siRNAs may gain access to the targeted DNA by the effects of RNA
Pol-II opening up the targeted region (4) for the siRNA/RITS/RdRP
complex to gain access (5) leading to gene silencing (6). While in
human cells the mode of action may be different (B). The antisense
strand (data not shown) of the promoter directed siRNA may be targeted
to a low level expressed non-coding transcript (1) subsequently
allowing the siRNA/DNMT3A/HDAC/Suv39H1 complex to localize to the
targeted chromatin (2) leading ultimately to histone methylation and
initial silencing of the targeted promoter (3). Alternatively, the
antisense strand of the siRNA may gain access to the targeted promoter
by the effects of RNA Polymerase II (RNA Pol-II) on the targeted
promoter (4) resulting in the recruitment of the
siRNA/DNMT3A/HDAC/Suv39H1 complex (5), leading to histone methylation
and silencing of the targeted chromatin (6).
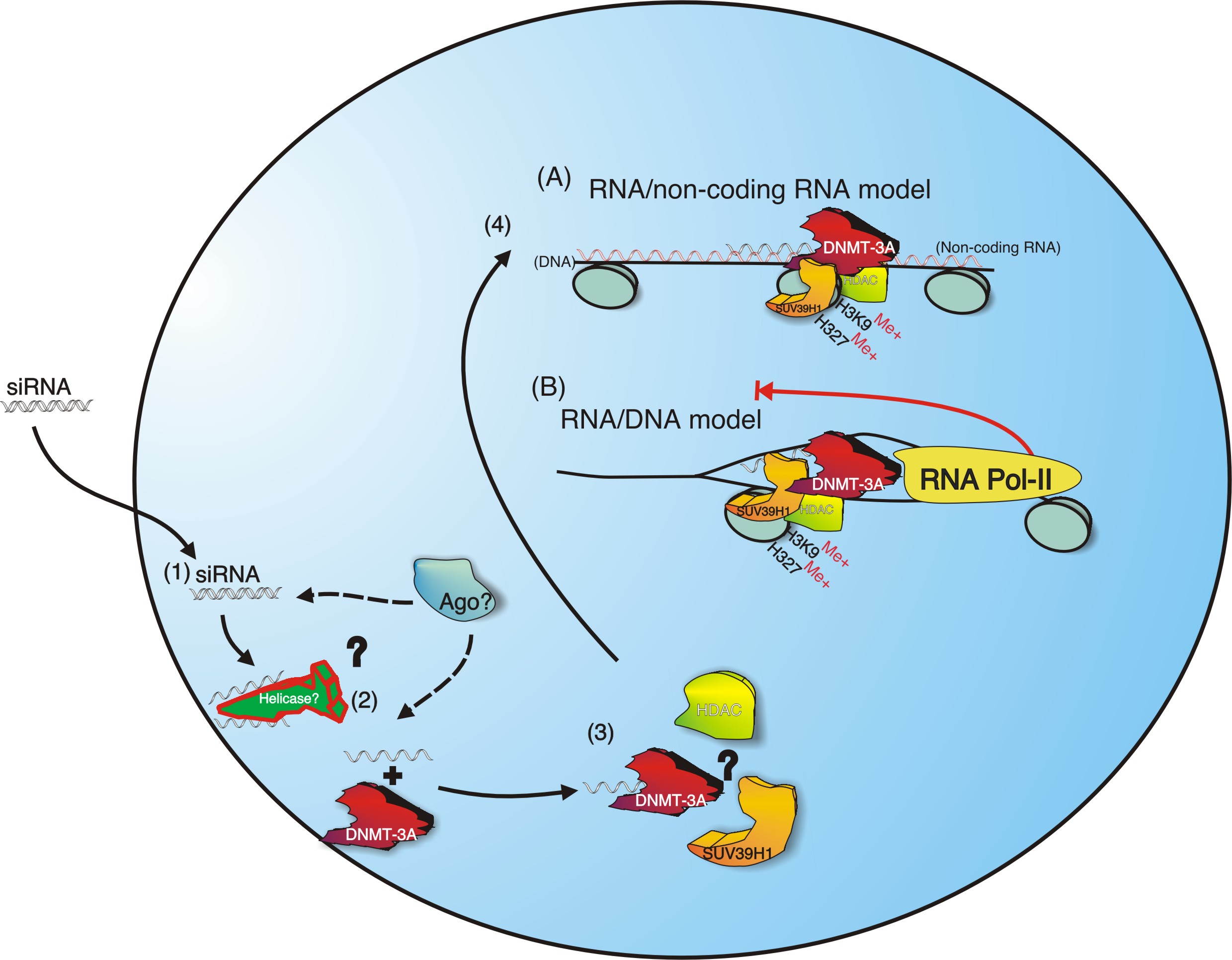
Model for
siRNA mediated TGS in Human Cells. SiRNAs are introduced by
nuclear specific MPG based transfection [51] into the target cells (1).
Once inside the nucleus the antisense strand of the siRNA (AS-siRNA) is
bound by DNMT3a (data not shown, (2)) (DNMT3b may also bind
siRNA)([85] and (K.V. data not shown)). Next the AS-siRNA/DNMT3a
complex may interact directly or already be bound by HDACs and/or
Suv39H1 [86, 87](3). The AS-siRNA probably then directs either the
AS-siRNA/DNMT3A complex with or without the HDACs and/or Suv39H1 to the
targeted promoter region, possibly via an interaction with a non-coding
transcript that is associated with the targeted chromatin (4) where
HDAC can deacetylate the respective histones (H3K9 and/or H3K27). The
deacetylation of H3K9 and H3K27 would then permit Suv39H1 to methylate
H3K9 and possibly H3K27 resulting in initial silencing of transcription
(5). If the silencing is re-enforced the gene may become methylated and
permanently silenced. It should also be noted that Argonaute 2 (Ago-2)
may also be involved in the observed promoter specific silencing
through the proposed pathway or an alternative yet to be described
pathway.